| GISdevelopment.net ---> AARS ---> ACRS 1995 ---> Forest/Vegetation Mapping |
Estimating Vegetation
Function from Satellite Data in Seasonal Tropical Environments
Kazue
Fujiwara
Yokohama National university
Institute for Environmental Science and Technology Hodogaya-ku,
Tokiwadai 156, Yokohama 240, Japan
Elgene 0. Box
Tokyo University, Institute of Industrial Science
Minato-ku, Roppongi 7-22-1, Tokyo 106, Japan 1
(permanently: University of Georgia,
Geography Department, I ~ Athens, Georgia 30602-2502, USA)
Abstract
Yokohama National university
Institute for Environmental Science and Technology Hodogaya-ku,
Tokiwadai 156, Yokohama 240, Japan
Elgene 0. Box
Tokyo University, Institute of Industrial Science
Minato-ku, Roppongi 7-22-1, Tokyo 106, Japan 1
(permanently: University of Georgia,
Geography Department, I ~ Athens, Georgia 30602-2502, USA)
Highly seasonal tropical environments have provided convenient situations for demonstrating the capabilities of satellite sensors to detect changes in the landscape, based on clear seasonal changes in the foliation status of the vegetation .Foliation status does not always vary so obviously, however, and even when it does, such changes do not necessarily translate directly into high or low levels of functional activity, such as , net primary production or net carbon flux (net ecosystem .production) .Southeast Asia is an especially good place to study : this problem, due to its full range of deciduous to largely .evergreen vegetation, all in a highly seasonal monsoon climate. Production, water budget, and overall net carbon flux of : different vegetation types are simulated using a climate-based model of vegetation and detrital metabolism. Parallel monthly results are shown for vegetation phenology, satellite data (NDVI) ; and simulated carbon balance. Such models of vegetation function .can and should be used for better calibration of satellite data .in various parts of the world.
Introduction
When rainfall falls to near zero for several months each year and 5 defoliation is relatively complete, satellite data can infer r changes in ecosystem function quite effectively (e.g. Tucker et al. 1985) .The main processes of interest include primary r production (photosynthesis plus assimilation of its products into new biomass) , autotrophic respiration (energy use for maintenance .and growth) , detrital decomposition, and the resulting net biosphere-atmosphere C02 flux. This overall net metabolic balance of the landscape (vegetation-detritus system) and sometimes even of the living vegetation (net production) normally J becomes negative during defoliation (dry season) .It can also J become negative, however, in green landscapes, during their growing season, if respiration is higher than photosynthetic r gains (Box et al. 1989) .As a result, in using satellite data to estimate biotic function, it seems wise to, look at, parallel estimates of vegetation metabolism, as are provided by simulation models based on field-measured metabolism and climatic data. Southeast Asia is a good place to study this problem, due to its full range of deciduous to evergreen but seasonal vegetation
Data and Methods
Thailand has remaining areas of moderately natural forest as well as larger areas of substitute landscapes covering the full range from evergreen to deciduous and from forest to open savanna (e.g. ~ Ogawa et al. 1961, Santisuk 1988) .The composition, structure "~"~ and productivity of the relatively natural vegetation have been ~ studied in the field for some time (e.g. Kira et al.1967, Smitinand 1980) .The main natural landscapes and their phenology are shown schematically in Figure 1 (from Fujiwara 1993)
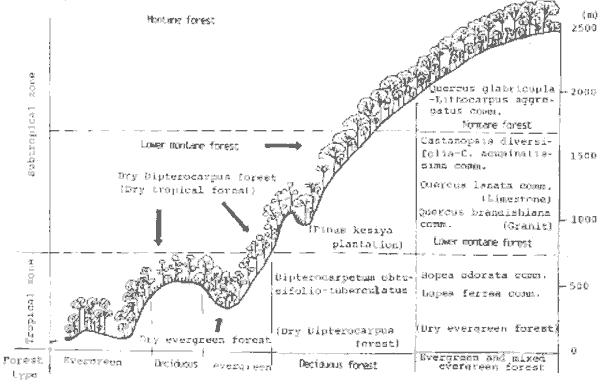
Figure 1. Schematic Zonation of Natural and some Substitute Landscapes (Vegetation) of Thailand.
Net primary production (NPP) is gross primary production (GPP, essentially photosynthesis) minus respiration (R) .GPP increases with warmth and water availability (other factors not limiting, Lieth & box 1977) , while respiration increases at least quasiexponentially (per unit biomass) with increasing temperature (e.g. Kira 1975) .NPP thus also increases with warmth and wetness (Lieth & Box 1972) but becomes a smaller fraction of GPP under warmer conditions (high respiration losses) and can become negative if GPP is low or zero, as in, an extreme dry season. Litterfall (dead leaves and other detritus) is also related to climatic conditions (Meentemeyer et al. 1982) ; detrital decomposition (D) also increases with warmth and wetness (Meentemeyer 1985) , and completes. the equation for overall net ecosystem production:
each component of which can be expressed in units of dry biomass or carbon equivalent. All components of this vegetation-detritus energy budget, as well as atmosphere-biosphere water fluxes, can be simulated at monthly intervals by the model MONTHLYC (Box 1988) , which is based on individual process models calibrated globally to climatic data and now NDVI from field-measured annual metabolic data. In the absence of field data for monthly metabolism, monthly climatic GPP is driven by AET (actual evapo- transpiration) , its closest apparent correlate (cf. Box et al. 1989) .Respiration is a function of temperature and is simulated therefrom using the usual Q1O value of two. For comparison purposes, field-measured values for natural and artificial primary production in Southeast Asia are shown in Table 1.
Satellite data for Southeast Asia, especially NDVI data, have been received, converted into imagery, and interpreted by various groups, including the Asian Institute of Technology in Bangkok, the Institute of Industrial Science at Tokyo University, and the NASA Goddard Space Flight Center in Washington. For this study, however, individual NDVI pixel values are used, since it is at pixels corresponding to field measurement sites that comparisons, calibrations and validations must ultimately be done.
| Standing Production | |||||
| Location | Vegetation | Biomass | Gross | Net | Source |
| Khao Chong | rainforest | 32.6 | 12320 | 2860 | (4) |
| Ping Kong | monsoon forest | 29.1 | 7000 | 1190 | (6) |
| savanna | woodland | 7.8 | 3200 | 760 | (6) |
| Pasoh | rainforest | 52.5 | 8190 | 2740 | (5) |
| Cheko | rainforest | 41.5 | 11700 | -- | (3) |
| Substitute/successional vegetation: | |||||
| Gorakhpur | teak plantation | 74.3 | -- | 2665 | (1) |
| Varanasi | succ. dry forest | 3.5 | -- | 301 | (1) |
| subst. grassland | 1.0 | 2166 | 1177 | (2) | |
Biomass values are in units of kg/m2, production values in units .of g/m2/year, each for dried biomass. Net production was generally estimated by harvest methods, whereas gross production ! involved gas-exchange, micrometeorological and other methods. Sources: 1 = Cannell 1982, 2 = Dwivedi 1971, 3 = Hozumi et al. 1969, 4 = Kira et al. 1967, 5 = Kira 1978, 6 = Yoda 1967.
Results
Parallel monthly values of NDVI, climate and correspondingly t driven simulated landscape metabolism are compared in Table 2 for Ping Kong, a site in northern Thailand {near Chiang Rai) where vegetation metabolism has been field-measured {Yoda 1967) .The ! r potential natural vegetation at Ping Kong is mixed deciduous ~ "monsoon" forest, but the actual landscape is evidently partly r evergreen, as seen from the NDVI values, which never drop near r zero. NDVI does parallel to some extent the seasonal course of precipitation and AET, all rising sharply in May. The monthly patterns of climate-driven and NDVI-driven GPP are quite different, however, and thus also the resulting monthly patterns of NPP and overall metabolic balance {equation 1) .Even with significant NDVI values and NDVI-simulated GPP during the dry season, however, the corresponding NPP still falls negative in April and the overall C02 balance is negative for five months {with a time lag due to the smaller amplitude of the GPP curve for the more evergreen actual landscape) .
Conclusion
Although time-consuming and difficult to measure, some field measurements of vegetation metabolism are available, in Southeast Asia and elsewhere. Based on these, it is possible to estimate seasonal patterns of vegetation function by simulation modeling. It is often especially difficult to interpret satellite data in humid/seasonally humid tropical areas, due to persistent cloud cover, a narrow range of NDVI variation, light-trapping effects of tall forests with dark green leaves, etc. NDVI data cannot directly estimate phenomena which become seasonally negative, including net primary production and overall C02 flux, since there is no way to calibrate the zero-point against NDVI values which are always positive. NDVI can be used to estimate gross production, but in order to get net production, a metabolic simulation model is still needed to estimate respiration. Such models of vegetation function can and should be used for better calibration of satellite data in various parts of the world.
References
- Box, E. 0. 1988. Estimating the seasonal carbon source-sink geography of a natural steady-state terrestrial biosphere. J. Appl. Meteorol. , 27: 1109-1124.
- Box, E. 0., B. N. Holben, and V. Kalb 1989. Accuracy of the AVHRR Vegetation Index as a predictor of biomass, primary productivity, and net C02 flux. Vegetation, 80:71-89.
- Cannell, M. G. R. 1982. World Forest Biomass and Primary Production Data. London: Academic Press. 391 pp.
- Dwivedi, R. 1971. Suitability of measuring carbon assimilation rates for evaluation of solar energy conservation and dry- matter production in plants. Trop. Ecology, 12:123-132.
- Fujiwara, K. 1993. Rehabilitation of Tropical Forests from ,is Countryside to Urban Areas. In: Rehabilitation of Tropical C Forest Ecosystems (H. Lieth & M. Lohman, eds.) , pp. 119- ~ 131. Amsterdam: Kluwer Acad. Publishers. ~
- Hozumi, K., K. Yoda, Sh. Kokawa and T. Kira 1969. Production ecology of tropical rainforests in southwestern Cambodia. -;8 I and II. Nat. Life BE Asia (Kyoto) , 6: 1-57, 57-81
- Kira, T., H. Ogawa, K. Yoda and K. Ogino 1967. Comparative : ecological studies on three main types of forest vegetation ~ in Thailand. Part IV: Dry-matter production, with special reference to the Khao Chong rainforest. Nat. Life BE Asia ,~ (Kyoto) , 6:149-174
- Kira, T. 1975. Primary production of forests In. Photosynthesis and Productivity in Different Environments (J. P. Cooper, ed.), pp. 5-40. IBM series, vol. 3. Cambridge j,;ii1, University Press.
- Kira, T. 19?8. Primary productivity of Pasoh forest synthesis. Malay. Nat. J., 30:291-297
- Lieth, H. and E. 0. Box 1972. Evapotranspiration and primary productivity; C. W. Thornthwaite Memorial Model. Publ. in Climatology (Uni v. of Delaware) , 25 (3) : 37-46.
- Lieth, H. , and E. 0. Box 1977. The gross primary productivity pattern of the land vegetation: a first attempt. Tropical E 1 18 109 115
- Meentemeyer, V. 1985. Climatic control of litter dynamics in tropical forest. In: Ecology and Resource Management in Tropics (K. Misra, ed. ) , pp. 302-310. Varanasi: Bhargava
- Meentemeyer, V., E. 0. Box, and R. Thompson 1982. World patterns and amounts of terrestrial plant litter production. BioScience, 32:125-128.
- Ogawa, H., K. Yoda, and T. Kira 1961. A preliminary survey on the vegetation of Thailand. Nat. Life BE Asia (Kyoto) , 1:22-157.
- Santisuk, T., 1988. An Account of the Vegetation of Northern Thailand. Wiesbaden: Franz-Steiner-Verlag. 101 pp.
- Smitinand, T. 1980. Thai Plant Names. Bangkok: Royal Forest Department. 379 pp.
- Tucker, C. J., J. R. G. Townshend, and T. E. Goff 1985. African land-cover classification using satellite data. Science,
- Yoda, K. 1967. Comparative ecological studies on three main , types of forest vegetation in Thailand. III. Community respiration. Nat. Life BE Asia (Kyoto) , 5:83-148.